And God made the beast of the earth after his kind,
and cattle after their kind,
and every thing that creepeth upon the earth after his kind:
and God saw that it was good.
And God said,
Let Us make man in Our image, after Our likeness:
and let them have dominion over the fish of the sea,
and over the fowl of the air,
and over the cattle,
and over all the earth,
and over every creeping thing that creepeth upon the earth.
(Genesis 1:25-26)
Human DNA is 98.4 percent identical to the DNA of chimps and bonobos, a lesser-known chimp-like ape.
That's the critical question," said de Waal, a renowned primatologist.
A more recent "Study found only 86.7% genetic similarity when segments of human and chimpanzee DNA (totaling 1,870,955 base pairs) were laid side by side. This study also included indels (insertions/deletions) in addition to substitutions." ref: Tatsuya Anzai st al., "Comparative Sequencing of Human and Chimpanzee MHC Class | Regions Unveils Insertions/Deletions As the Major Path to Genomic Divergence," Proceedings of the National Academy of Sciences, USA 100 (2003); 7708-13
| Human | 100 % |
|---|---|
| Chimpanzee | 86.7 % |
| Nematode | 75 % |
That puts the Chimpanzee at
LESS than HALFWAY
between a Worm and Human!
And even more recently researchers found that about
80% of the proteins in the
human
and chimpanzee genomes are different.
(reference: Galina Glazko, Vamsi Veeramachaneni, Masatoshi Nei and Wojciech Makalowski, "Eighty Percent of Proteins are Different between Humans and Chimpanzees," Gene volume 346 14 February 2005, Pages 215-219 )
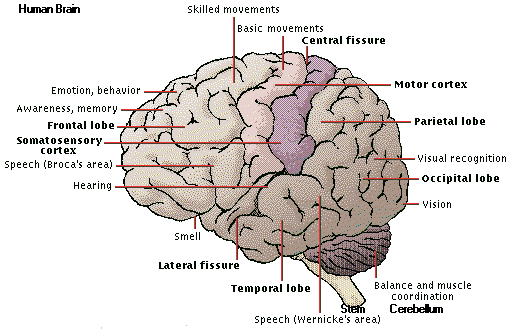
Language is one critical difference. Humans are "left brained" -- the planum temporale that controls language is
much larger on the left. (Laymen may know the term because the left brain also controls right-handedness.)
Human infants are born as language learning machines.
"First, infants begin forming up categories for the common speech sounds they hear,
not whole words so much as the little units we call phonemes, less than a tenth
of a second in duration. Categories allow them to generalize across speakers,
so that the mothers /ba/ sound and the fathers somewhat deeper /ba/ sound are
treated the same despite their differences. By about a year of age, babies stop
hearing many of these differences, having standardized them.
By a year of age, babies are discovering patterns in the strings of phonemes and
acquiring six to nine new words every day, just from the examples they hear
(long before they begin speaking them). The words acquiring meaning, the phonemes
remain meaningless. You can say kids are like sponges soaking up words but
thats too passive a notion, one of the reasons I prefer the more active
acquisitive as the characterization.
So kids have pyramided words atop the phonemes, and now have compound structures
made from building blocks. But then they do it again, discovering patterns in
the strings of words they hear and inferring the grammar of that particular language:
ways of making plurals and past tenses and nested phrases. This happens between
the ages of 18 and 36 months. Then theyre off detecting patterns on even longer
time scales, that of the collection of sentences we call a story. They infer that
a satisfying story has a beginning, middle, and a wrap-up ending - and then they
start demanding proper endings for their bedtime stories." (from http://faculty.washington.edu/wcalvin/2000/SanDiegoSpeech.htm)


Another great and obvious difference is brain size and function. From our brain comes control of the involuntary and voluntary functions, the sensory perceptions, memory, emotions, consciousness, and intelligence.
Endocranial Volume (in cm3)
[From
Aiello and Dean 1990, Pp. 193]
| Species | Lower 95% Limits for Mean | Upper 95% Limits for Mean |
Mean |
|
Human |
1,159 |
1,243 |
1,201 |
|
Chimpanzee |
391 |
409 |
400 |
|
Gorilla |
452 |
486 |
469 |
|
Orang |
385 |
409 |
397 |
|
|
"Among mammals there are two patterns of brain growth. The first pattern is called altriciality. In this pattern the animal is born helpless and extremely immature. The brains of altricial animals are usually half the size of the adult's, and double in size by adulthood. Because of this it takes lots of parental effort to raise the young. Animals following this pattern usually have litters and perform this care for multiple offspring at once. Cats, with their blind and helpless kittens are altricial. The other pattern is precocial. In this pattern the offspring are usually born single and from birth are able to get around quite well. Their brains are nearly adult size at birth. The are alert and all their organs are functioning. An example of this pattern is the horse, the wildebeest etc., where the young will run with the herds within minutes.
Now, according to Walker and Shipman (1996, pp220-222), altricial species almost never have bigger brains than precocial species. The reason is that for all mammals save one, the brain grows rapidly during gestation but then grows less rapidly after birth. There is a kink in the graph of brain size vs. time which occurs at birth. Altricial species whose immature state at birth and subsequent slow down in the rate of growth forever remain behind the more maturely born precocial species.
What humans seem to have accomplished is the trick of keeping the brain growing at the embryonic rate for one year after birth. Effectively, if humans are a fundamentally precocial species, our gestation is (or should be) 21 months. However, no mother could possibly pass a year old baby's head through the birth canal. Thus, human babies are born "early" to avoid the death of the mother. Walker and Shipman (1996, p. 222) write:
"Humans are simply born too early in their development, at the time when their heads will still fit through their mothers' birth canals. As babies' brains grow, during this extrauterine year of fetal life, so do their bodies. About the time of the infant's first birthday, the period of fetal brain growth terminates, coinciding with the beginnings of speech and the mastery of erect posture and bipedal walking."
This pattern of growth has huge implications. Every other primate doubles their brain weight from birth to adulthood. But due to the early birth of humans, we triple our brain's birth rate. Our last 12 month of fetal growth rate of the brain occurs outside the sensorially deprived womb. The vast quantities of sensory input during the first year of life affects the rate and nature of the neural connections. Because of this year of helplessness, parents must provide close physical and emotional support for the infant. Unlike chimp babies who can cling to their mother's fur, human infants cannot even hang on to mother in spite of having the hand reflex. The mother has no fur because she sweats and she sweats because of a big brain which is why she gives birth to her child early. This early birth then requires the mother to care for the infant and increases the bond between mother and child which partially makes us human." (from http://home.entouch.net/ dmd/sweat.htm)
The physical differences are many!
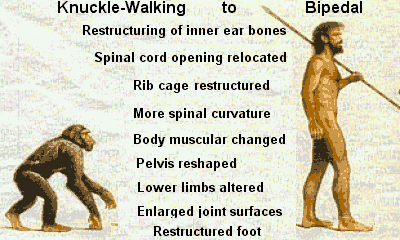
The structural changes to the feet are not simple in that they involve an elaborate plantar aponeurosis, strong plantar ligaments, longitudinal arches, an enlarged musculus flexor accessories, a remolded calcaneocuboid joint, a long tarsus, reorganization of the neuronal innervation and blood vascularization, an adducted (non-opposable) hallux and shortened toes. Many are quick to point out that primates needed to possess opposable digits so they could grasp, and quickly swing from tree branches. But then, how about the feet, did they devolve, humans have the opposable thumb, but not the opposable big toe?
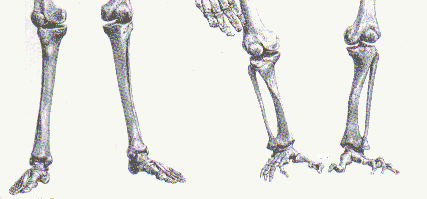
In his 1999 paper published in the Journal of American Podiatric Medical Association, R. Kidd described the evolution of the feet as follows:
"The evolution of the human foot presents an obfuscation: explanations for its occurrence and the exact nature of the mechanisms of change are still not fully understood."
For more details on the differences between humans and the other primates see Appendix A & B."Richard Klein, professor of anthropological sciences, in his new book, The Dawn of Human Culture (John Wiley & Sons, April 2002), proposes a plausible theory for the latest stages of human evolution. "It's a nontestable hypothesis," Klein admits. "The book is about human evolution as I understand the record. My genetic explanation for the major behavioral change 50,000 years ago is the most plausible one, but I can't prove it." For Klein this neural mutation hypothesis is the most economical explanation of why anatomy and human behavior drifted apart. Fossilized skulls reveal little about the brain underneath. But a gene mutation may have changed critical neural processes such as speech and language.
The now widely accepted "Out-of-Africa-2" hypothesis is based on the appearance of anatomically and behaviorally modern humans on a small patch in Eastern Africa as recently as 50,000 years ago. All of a sudden these early modern humans developed a new repertoire of hunting skills, novel forms of social interaction and a sense of art. They became creative innovators expanding their mental and technical capabilities. These new achievements drove the early modern humans out of Africa to spread over Europe and Asia. Within a short period of only about 15,000 years they supplanted the Neanderthals in Europe and other nonmodern humans in other parts of the world.
The cause for the drastic change in behavior in the early modern
humans is unknown. But the most plausible explanation for the
success of modern humans is a sudden biological change.
Some of the new (and old) evidence is ambiguous, circumstantial, or even contradictory, but this is inevitable in historical science, which has more in common with a criminal trial than it does with a physics experiment. " (examples, Toumai, Monkey or man?)
(Stanford Report, April 26, 2002, "Anthropologist explores the dawn of human culture in new book" by Christian Heuss and "The Dawn of Human Culture" by Richard G. Klein with Blake Edgar, 2002)
"A fortuitous mutation" , the basis for the entire "theory of evolution", multiplied over and over, from organic slime to the modern human!! "It's a nontestable hypothesis", doesn't this also apply to and describe the entire "theory of evolution"?
It is doubtful that we will ever fully understand the human brain. Some want to say it is no more than a wonderful organic computer, but that seems like a great over simplification. It is a computer that can change and modify its functionality. Tests with people who have had brain surgery and lost some capabilities indicates that over time part or much of the lost functionality can be recovered with non- damaged portions of the brain assuming capability that lost brain sections once controlled. It has "plasticity", it is continually changing. These changes come about by synapses becoming activated or deactivated through the growth or contraction of dendritic spines. And these changes can take place in surprisingly short times. Varying genetic factors combined with varing environmental factors and the "plasticity" of the brain gives us "individuality".
Domination; man was charged to subdue the earth and have dominion over all the life forms that occupy it (Genesis 1:28). Many scientists tell us that the other primates preceded humans by many millions of years, and if this was true, then they had their chance to dominate. And obviously they failed to do so. To our knowledge there is no evidence that either Cro-Magnon man or Neandertal qualified since there is no apparent evidence that they had dominion. Some will argue that presently humans are royally messing it up, and possibly they have a good point. For as one looks around at the state of the human society at this time, one can see many things that seem to be rather undesirable, and which would seem to be detrimental to the ecology.
Conclusion: Question: Do we understand the full implications of "in Our image, after Our likeness"?
Answer: Obviously not!!
But this we do know, the differences between humans and the other primates is wonderfully great!
Only man has what it takes to "have dominion" !!


Appendix A: Physical Primate differences
(from:http://www.dla.utexas.edu/depts/anthro/people/faculty/cbramblett/ant301/seven.html)
Human Skeletal and Dental Anatomy
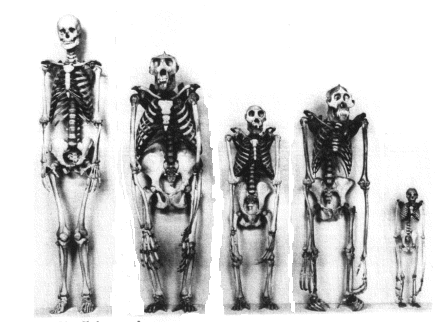
Humans are contrasted with chimpanzees to
illustrate the unique features of our anatomy. Other primate species are
included when they are needed to demonstrate a point.
Primates have four types of teeth - incisors, canines, premolars and molars. Incisors are like tiny shovels or spatulas that cut food. Canines are generally pointed, stabbing teeth that can serve as weapons in most primates. Premolars and molars have large crown surfaces that shear and crush food during mastication.
The number and type of teeth are summarized by
listing only elements on one side from incisor, canine, premolar to molar
(mesial to distal) for each jaw. Thus the primitive mammalian formula of
represents a hypothetical mammalian ancestor with 44 teeth.
Whatever the ancestral condition, no living primate has more
than three incisors or three premolars on each side in either maxilla or
mandible. Prosimians and Platyrrhines have three premolars; Catarrhines have
only two. Third molars are often absent in some primate genera.
Once a tooth is lost in a species, it is usually not reproduced again. Thus
ancestral forms generally may have more teeth, but not fewer teeth of a
particular kind than their descendants.
A tooth consists of a crown, the portion covered
with enamel, and a root of dentine covered with cementum. The
interior of the tooth is the pulp chamber that contains soft cellular tissue.
The primary mineral in both bone and dental structures is crystals of apatite
- a form of calcium phosphate. Dentine is a bone-like substance (about 75%
mineral) but enamel is much more heavily mineralized (96% mineral by weight).
Cementum, whose composition differs only slightly from that of dentine,
attaches the tooth to its periodontal ligament and provides a dynamic
interface between tooth and surrounding bone, while enamel forms a crystalline
cap over the working surface of the crown.
Primates have two sets of teeth during their lives, a deciduous set that is replaced by permanent teeth during childhood and adolescence. The human dental formulas are:
| Deciduous | Permanent | |
Maxilla |
2 1 2 |
2 1 2 3 |
Mandible |
2 1 2 |
2 1 2 3 |
Note that humans have no deciduous molars.
Substantial individual variation occurs in tooth number, but most often the
variant is the loss of a tooth at the boundary between tooth kind
(incisor/canine/premolar/molar). Infrequently there can be supernumerary teeth
or additional teeth near a boundary. In traditional notation the most mesial
permanent premolars and the most distal incisors are lost in primates.
Consequently the premolars of primates are designated the second, third, and
fourth premolars -- presumably homologous to the primitive mammalian second,
third, and fourth premolars. Humans are also missing fourth premolars, leaving
us with premolars three and four. For example, the notation pm3 refers to
mandibular third premolar, the premolar next to the canine on the lower
jaw.
The greatest differences between humans and chimpanzees occur in the canine teeth. Small peg-like human canines do not project from the tooth row. In contrast, chimpanzee canines are much larger, robust, and project far above their tooth row. Diastemas, gaps in the tooth row of the maxilla allow projecting mandibular canines to pass the opposing canine and incisor during occlusion. The maxillary canine passes the buccal side of its opposing pm3, allowing the lingual surface of the canine to make contact with a blade-like sectorial surface on the premolar. Humans lack the large diastema and the human pm3 is non-sectorial. Human anterior teeth (canines and incisors) are greatly reduced in size and human incisors are positioned close to a transverse plane that passes through the canine teeth. Chimpanzee incisors are positioned well forward of this plane. Consequently the parabolic or elliptical human dental arcade contrasts sharply with the U-shaped arcade of chimpanzees. Human molars tend to be rounder and more compact than chimpanzee molars. Occlusal molar surfaces of human teeth are relatively flat, and quickly become even flatter with attrition
The skull consists of 28 bones that are
conveniently described as the bones of the calvarium (supporting and
surrounding the brain) and those of the face. Tiny bones of the middle ear,
the conchae and vomer in the nose, and the ethmoid of the orbital vault are
not easily seen. Although usually not considered part of the face, the hyoid
bones form a skeletal element for the larynx. In addition to the openings that
represent the eyes, nose, mouth and ears, the skull has numerous foramina for
the passage of nerves and vessels. The bones of the cranium (the skull
minus the mandible) are joined by irregular sutures that are
obliterated after growth ceases.
Seven major differences between humans and
chimpanzee skulls include:
(1) brain volume -- The human skull has a three
-fold greater endocranial volume - reflecting a larger brain
size, about 1200 cc in human and about 400 cc in chimpanzees. The larger size
of the human calvarium allows ample surface area for the attachment of
the powerful M. temporalis. Fibers from this muscle attach to the skull in the
fossa temporalis and pass behind the zygomatic arch to insert on the
coronoid process and anterior margin of the ascending ramus of
the mandible. The temporal lines or temporal crests on the
calvarium mark the terminal fibers of M. temporalis. If brain volume is small,
producing a small calvarium (as in a chimpanzee), there may not be
enough surface area on the calvarium for M. temporalis fibers to attach. Where
fibers from the opposite muscle meet, a bony crest, in this case a sagittal
crest is formed. Crest formation usually signifies a small calvarium relative
to muscle size.
Endocranial Volume (in cm3)
[From
Aiello and Dean 1990, Pp. 193]
| Species | Lower 95% Limits for Mean | Upper 95% Limits for Mean |
Mean |
|
Human |
1,159 |
1,243 |
1,201 |
|
Chimpanzee |
391 |
409 |
400 |
|
Gorilla |
452 |
486 |
469 |
|
Orang |
385 |
409 |
397 |
The larger human brain volume also means that the calvarium is large relative to the size of the face. As the brain expands (comparing chimp and human), it appear to flex on an axis around the pituitary fossa. This produces flexion of the cranial base, a downwards shift in the posterior part of the cranium, and a forward rotation of the foramen magnum.
Expansion of the occipital lobes and cerebellum balloons the posterior cranial fossa, that in turn correlates typically with an asymmetrical cranial venous sinus system. In the human, a superior sagittal sinus drains venous blood in a transverse sigmoid route to the internal jugular veins. Enlarged occipitomarginal sinus systems typical of humans are infrequent in chimpanzees. The human middle cranial fossa expands with enlarged temporal lobes of the brain expanding the mid-section of the calvarium outward above the petrous portion of the temporal bone and the glenoid fossa. In chimpanzees, the calvarium is narrower than the cranial base but the human calvaria has its maximum width high on the parietal eminences rather than low at the cranial base. The anterior cranial fossa expands with the human frontal lobes. Consequently humans lack the postorbital constriction that occurs in chimpanzees and humans exhibit frontal eminence (forehead) above a greatly less obvious supraorbital torus (eyebrow ridge). Olfactory nerve tracts pass through the cribriform plate, a sieve-like structure in the middle of the ethmoid bone. In humans the median plane of the cribriform plate is the site of a process, the crista galli. This process is diminutive or absent in chimpanzees.
Ballooning of the calvaria in the temporal and occipital areas associated with the expansion of brain volume produces an apparent forward shift in the relative position of the foramen magnum. The human foramen magnum lies on a line, the bitympanic line, that connects the most inferior points on the lateral end of the right and left tympanic plates. The chimpanzee foramen magnum lies well behind the bitympanic line.
(2) nuchal crests -- Muscles that attach to the cranial base serve to position, move, and stabilize the head and cervical vertebrae. From a functional perspective, the skull is also the most superior point of origin for muscles (such as M. trapezius) that provide movement and stability to the back and shoulder. Consequently, some of the powerful muscles that attach to the skull have large attachment surfaces. The human occipital is a cup-like bone with a noticeable eminence, the external occipital protuberance. It is marked by nuchal lines and markings that represent attachment surfaces. The nuchal crests of the chimpanzee are more extreme. Its small skull combined with a much more robust musculature produces strong crests. Due to the flexion of the cranial base with expanded endocranial volume, the superior nuchal line is lower on the human skull.
(3) mastoid process -- The human mastoid process, the attachment surface of the sternocleidomastoid muscles, is distinct and separated from the outline of the occiput by a digastric fossa.
(4) premaxilla -- The smaller human incisivum, a homologue of the chimpanzee premaxilla, unites with the maxilla near the time of birth. The chimpanzee premaxilla is expanded to support the large and forward projecting incisors. It unites with the maxilla postnatally and the suture between it and the maxilla becomes obliterated.
(5) facial prognathism -- The projection of the face beyond the calvaria is greater in chimpanzees than humans. Flat faces are called orthognathic and projecting faces are called prognathic. The craniofacial angle, the angle between the most anterior point on the maxilla, the most anterior point of the sphenoid bone and the most anterior point of the foramen magnum, is used to quantify the extent to which the face projects beyond the neurocranium. Since this angle can not readily be measured on intact skulls, the angle that a sellion-prosthion line makes with the Frankfort plane serves as a convenient measure of facial projection. [The "Frankfurt plane" is the plane that passes through porion (right and left) and orbitale (right and left).] The porion is the most lateral and superior point of the external auditory meatus. The orbitale is the lowest point on the infraorbital margin. The sellion is the deepest point in the hollow beneath the glabella in the median plane. The glabella is the most anterior point in the median plane on the supraorbital torus. The prosthion is the most anterior point on the maxilla in the median plane.]
(6) chin -- The human mandible is reinforced by a bar of bone, the mental protuberance, that strengthens the symphysis, the union of right and left halves of the bone. The lingual or posterior surface of the symphysis bears a pair of genial tubercles that represent muscle attachment sites for M. genioglossus and M. geniohyoid. The ape mandible lacks a mental protuberance and is reinforced by an inferior transverse torus or "simian shelf". Viewed from above, the contrasting shape of the dental arcades is distinct.
(7) hyoid -- The human hyoid is a U-shaped bone just above the larynx. The stylohyoid ligament attaches the lesser horn of the hyoid to the styloid process of the temporal bone. Generally, these styloid processes point to the lesser horns of the hyoid bone. The chimpanzee hyoid is expanded anteriorly to accommodate a laryngeal air sack, and is located higher in the neck
The human vertebral column consists of 33 vertebrae divided into five functional regions.
1. Seven cervical vertebrae, easily recognized by their transverse foramina, form the skeleton of our neck. The joints (articular surfaces) between cervical vertebrae are very mobile.
2. Twelve thoracic vertebrae, mobile only in the coronal plane, support the ribs.
3. Five robust lumbar vertebrae in the lower back are tightly articulated to withstand the weight of the torso.
4. Five sacral vertebrae fuse to form the sacrum, the component of the axial skeleton in the pelvis.
5. Four caudal vertebrae extend downward from the sacrum. In adults these caudal vertebrae unit to form the coccyx, a hidden human tail that partially blocks the inferior pelvic aperture.
Viewed from the side, humans have a series of four curvatures. The dorsal outline of the cervical and lumbar regions are concave while the thoracic and sacral regions are convex. The forward curve of the lumbar region is called lordosis and that of the thoracic region is called kyphosis.
Chimpanzees generally have one more thoracic, one less lumbar vertebra, and one less caudal vertebra than do humans. Both normally have seven cervical vertebrae and normally the combined thoracic, lumbar, and sacral regions consists of 22 vertebrae. Chimpanzees lack the extreme curves of the human column, and the angle between the lumbar and sacral region is more acute .
Average Number and Range of Number of Vertebrae in Humans and Chimpanzees (after Schultz 1961)
Species Thoracic Lumbar Sacral TLS Caudal Human Mean 12.0 5.0 5.2 22.2 4.0
Chimpanzee Mean13.2 3.6 5.7 22.5 3.3 Human Range 11-13 4-6 4-7 2-5 Chimpanzee Range 12-14 3-4 4-8 2-5 Thorax
The human chest consists of 12 paired ribs that articulate with the vertebral column. Ribs I through VII attach ventrally to the sternum. Ribs VIII through X terminate in cartilage extensions that eventually fasten to the sternum. Ribs XI and XII have free distal ends. The sternum is composed of six flat bones that fuse in adults to three units, the manubrium (segment I), body (segments II through V), and xiphoid process (segment VI). In some persons, especially in old age, the three units of the sternum may fuse to each other.
Consistent with their extra thoracic vertebra, chimpanzees usually have one extra rib (13 pairs). The human rib cage is slightly broader for its depth than the chimpanzee and the human thorax is less funnel-shaped. Chimpanzee ribs are also somewhat rounder in cross-section than human ribs
The shoulder is formed by the humerus, clavicle and scapula. The clavicle attaches firmly to the manubrium and acts as a strut to hold the shoulder joint away from the chest. Distally, the clavicle articulates with the acromion process of the scapula, a large triangular flat bone in the back of the shoulder. The glenoid cavity of the scapula articulates with the head of the humerus.
The most striking difference between the chimpanzee and human shoulder is in the proportions of the scapula. Human arm strength, much less powerful in movements when in a raised position, is reflected in the shape of the scapula that provides attachment surfaces and lever arms for muscles
The upper arm is a single bone, the humerus. The forearm is formed medially by the ulna, which articulates firmly by a hinge joint to the humerus, and laterally by the radius which is firmly attached to the hand. The radius pivots on the humerus and ulna to supply hand rotation (pronation) and is attached to the ulna by powerful interossesous muscles and ligaments.
The head of the humerus is useful in identification of gender of a mature unknown cadaver. If the maximum diameter of the head is greater than 45 mm, the individual is probably male. If the head diameter is less that 42 mm, it is probably female.
The chimpanzee distal humerus contrasts with the human. The human lacks the robust lateral supracondylar ridge, a high and robust lateral epicondyle, and the steep, sharp, lateral margin of the olecranon fossa . The chimpanzee forearm is relatively long in comparison to humans . Chimp radius and ulna are more curved than in humans and the chimpanzee distal radius has a radiocarpal joint surface that diverges medially. The major differences between human and chimpanzee limbs are contrasts in relative proportion. Chimpanzees have large powerful arms, slightly longer than their very short legs. Human arms are about 70% as long as human legs.
Long bone indices of Humans and Chimpanzees
[ From Aiello and Dean 1990, Pp. 249]
Species Intermembral Index Humerofemoral Index Brachial Index Crural Index Human (male) 69.7 71.4 77.9 82.4 Human (female) 68.5 69.8 77.0 81.3 Chimpanzee (male) 108.0 101.1 91.9 79.8 Chimpanzee (female) 109.4 102 92.4 80.4 Pygmy chimpanzee (male & female) 102.2 98.0 91.9 82.6
where:
Intermembral index = [(humerus + radius) x 100]/)femur + tibia)
Humerofemoral index = (humerus x 100)/femur
Brachial index = (radius x 100)/humerus
Crural index = (tibia x 100)/femur
The hand has three skeletal regions: (1) The carpus, a series of eight carpals, form the wrist. (2) The hand consists of five metacarpal bones. (3) Phalanges form the skeleton of the fingers. The thumb, digit I has two phalanges (proximal & distal), while digits II through V have three phalanges (proximal, middle and distal).
The chimpanzee hand is notably different from the human hand in the relative length of its digits. The chimp thumb is much shorter than in humans, phalanges exhibit much more robust insertion areas for flexor tendons, and metacarpals have transverse ridges across their heads that limit dorsiflexion
The pelvic girdle is formed by the sacrum, coccyx, and the two coxae. Each coxa is attached by strong ligaments to the sacrum and to each other at the pubic symphysis. A coxa is formed by the fusion of three bones, the ilium, ischium, and pubis, which meet in the acetabulum or hip socket.
The human female has a larger birth canal than does a male. Consequently there is a constellation of characters that can be used to identify the gender of an unknown pelvis. The pelvic inlet of females is larger and has a greater absolute circumference. The superior ramus of the pubic bone is longer, increasing the pubic/ischium ratio. The greater sciatic notch is wider and forms a longer angle. The increased pubic length and laterally displaced ischia result in a wider subpubic angle. The growth and remodeling of the pubis produces extra bone at the symphysis, leaving a concave inferior ramus, a ventral arc that represents a previous border of the symphysis, and a narrow inferior pubic ramus. The female pubic symphysis is likely to be longer in its superior-inferior diameter and smaller in its dorsal-ventral diameters than is that of a male. Females are more likely to have a well-developed preauricular sulcus, and those who have borne children may have pits or guttering along the dorsal border of the pubic symphysis. Since they have smaller femurs, females have smaller acetabula.
Sexual Dimorphism in the Primate Pelvis
(After Schultz 1949 and A & D 1990
Species n Mean Min Max Human (male) 50 79.9 71.0 88.0 Human (female) 50 95.0 84.0 106.0 Chimpanzee (male) 21 86.2 69.7 95.2 Chimpanzee (female) 30 87 78.9 98.9
Differences in the pelvis reflect the differences between the habitual bipedal locomotion of humans and quadrupedal movements of chimps . The pelvis of the two primates have radically different form and locomotor function. The relative width of the iliac blade (width/length x 100) is much larger in humans (125.5) than in chimpanzees (66.0). The human ilium is broad and low, while the chimp ilium is narrow and high. The human has a barely noticeable iliac pillar or thickening which extends from the iliac tubercle to the acetabulum. The human has an anterior inferior iliac spine. The human acetabulum is larger, reflecting the larger head of the femur, and the superior border of the acetabulum is reinforced to sustain the pressure of bipedal walking. The inferior border of the ilium near the auricular surface forms a greater sciatic notch in humans. The auricular surface is larger in the human. The ape sacrum is long and narrow
The femur, the bone in the thigh, articulates ball and socket fashion in the acetabulum of the coxa. The lower leg contains a large medial bone, the tibia, which articulates with the femoral condyles to form the knee. Lateral to the tibia, the fibula is a small, irregular bone that provides attachment surfaces for muscles. Projections on the distal ends of tibia and fibula, the medial and lateral malleolus, form a secure slotted proximal structure for the ankle joint.
As in the head of the humerus, a maximum diameter of 45mm or greater for the femur head indicates a male gender.
The human femur is longer than the chimp femur and usually has an elevated pilaster that supports the linea aspera down the shaft. The angles of the head, shaft, and condyles contrast markedly with those of the ape. The load axis never intersects the shaft in a chimpanzee femur. The femoral condyles of the human are larger and more elliptical than are those of the chimp. The human femur has a larger articular surface and mid-shaft circumference when compared to the arm than does the chimpanzee. The chimpanzee has a similar ratio between femur length and tibia length but the chimpanzee leg (including both femur and tibia length) is much shorter relative to the arm. The chimpanzee proximal tibia is smaller, less well supported by the shaft, and has condyles than are more convex than is usual in humans
Like the hand, the foot has three anatomical regions: (1) The seven bones of the tarsus form the ankle and proximal half of the foot. (2) The middle portion of the foot consists of five metatarsals. (3) Phalanges, the skeletal elements of the toes, have the same number and arrangement as in the fingers.
The primary difference between the human and chimp foot is the contrast between prehensile functions of the ape foot and the anatomy of bipedal striding in the human. The chimpanzee foot has an opposable hallux and long phalanges. The human foot has greatly reduced digits, with all metatarsals parallel and an increase in the lever arm of the tarsus for striding. In the human foot, a longitudinal arch provides a shock absorbing and weight distribution system. The orientation of the ankle joint allows the tibia to take a straighter path over the foot during walking
The bones of the limbs and vertebral column are endochondral, that is, they are first formed as cartilage that is gradually replaced with bone. Centers of ossification in the cartilage gradually enlarge to form the bone. The body or shaft in long bones is called the diaphysis, or primary center of ossification. The ends of long bones have secondary centers of ossification, called epiphyses, that are separated from the diaphysis by cartilage plates. Eventually, the cartilage plates, the epiphyseal cartilage, are replaced by bone and the epiphyses unite with the diaphysis to form a single bone. The flat bones of the skull, mandible, and clavicles are intramembranous bones, formed from membranes, and generally do not have epiphyses
Much of primate anatomy reflects habits of movement and commonly utilized substrate. Since the powerful legs of most primates are slightly longer than their arms, the pelvis is normally higher than the head when standing quadrupedally. However a few species exhibit extreme locomotor specialization, emphasizing arms for arm-swinging, legs for leaping, or arms and legs comparable in length for quadrupedal climbing or walking on the ground. Thus, limb length (Intermembral Index or IM = Length of Humerus + Radius x 100/ Length of Femur + Tibia) can serve as an index of the relative emphasis upon the arm versus the leg for propulsion:
1. Short limbs with leg and arm comparable - quadrupedal and arboreal
These animals walk on larger horizontal tree branches as if they were pathways. Short, usually robust, arms and legs lower their center of gravity. IM index is usually about 80 but approaches 100 in howler monkeys. Other characteristics include moderately sized fingers and toes; very prehensile hands and feet; and relatively mobile shoulder joints located and directed sternally on the thorax. Some species, especially those which perform spectacular leaping feats, have flexible, elongated backs (with extra vertebra) and powerful musculature associated with the back and hind limb. A few species in the Americas have somewhat prehensile tails that serve to anchor the animal when it feeds near the ends of branches.
2. Long limbs with leg and arm of equal length - quadrupedal and terrestrial
Terrestrial quadrupeds tend to have shortened digits and elongated, robust tarsal and metatarsal elements. IM index is above 90. The shoulder joint, lying alongside the narrow and laterally flattened thorax, is oriented toward the ground. The weak clavicular-sternal joint is easily dislocated from the segmented sternum. Arms and legs, modified for powerful sagittal motions, have a relatively small range of movement. The humerus bears a prominent deltoid process (the attachment surface for the M. deltoideus) and the ulna has a large olecranon process, the insertion point of the M. triceps brachii, a powerful forearm extensor. Hands are pronated when in contact with the ground. One species, the patas monkey, is digitigrade (like a cat), with specialized anatomy for terrestrial running in which only fingers and toes make contact with the ground.
3. Very long limbs with leg and arm of comparable length - quadrupedal and arboreal with an emphasis on quadrupedal climbing and suspension
A few primate species in the Family Lorisidae combine quadrupedal suspensory climbing with quadrupedal arborealism, requiring great joint mobility and wide range of movement. IM index is about 90. Their hands and feet are particularly prehensile.
4. Arm longer than leg - brachiation and arboreal
Brachiation (arm swinging) is a special form of locomotion in which the body is suspended below branches. It allows utilization of small branches near the fringe of a tree canopy since the brachiator is suspended beneath its handholds. In contrast, a large bodied quadruped that tries to walk on a small branch has difficulty balancing as the supporting tree limb bends. A brachiator can easily exploit the very fringe of a tree canopy by dispersing its weight to the ends of several branches. New World brachiators use their prehensile tail as a fifth prehensile limb to further disperse weight. Most rapid brachiation is attained by using gravity to convert vertical height to speed. IM index is 100 or above.
Brachiation generally is associated with major alterations in the arm, hand, and thorax. The shoulder joint is positioned laterally and cranially on a barrel-shaped thorax. Robust muscles attach to the sternum, vertebral column, head, and rib cage, stabilizing the shoulder. The more powerful the arm movements, the more robust the stabilizing musculature must be. The clavicle acts a strut to stabilize the shoulder joint against a sternum whose segments unite to form a single bone. This clavicular-sternal joint is very strong and is not easily dislocated. A relatively round head of the humerus reflects a very wide range of motion. Additional elbow strength results from a more distinct separation of the radius and ulna on the articular surfaces of the distal humerus. The olecranon process of the ulna is small, allowing full extension of robust forearms. Brachiators tend to have reduced thumbs. If a thumb is present, it is folded out of the way against the palm where it does not interfere with elongated fingers that hook or snag handholds. The lumbar region of the vertebral column is shortened and stabilized, and a very mobile hip joint allows the foot to grasp anchorage in a wide range of positions.
There are several types of brachiators. Gibbons and siamangs, who use arm swinging as a major means of travel, are the best brachiators. Chimpanzee, gorillas, and humans are capable of this type of brachiation, but do not practice it as a primary means of locomotion. The orangutan combines quadrupedal climbing and brachiation, but like chimpanzees and gorillas, is typically a terrestrial quadruped.
At least one New World primate, the spider monkey, practices a variation of brachiation in which the body is kept vertical while brachiation is performed by hands, feet and sometimes the tail. This requires unusually long legs and mobile hips. When moving at slower speeds or while feeding, the spider monkey behaves as an arboreal quadruped. Its tail is the most prehensile of any primate.
5. Arm longer than leg - quadrupedal knuckle-walking and fist walking
Knuckle-walking is quadrupedal locomotion with the hands pronated and fingers flexed resulting in dorsal surfaces of the middle phalanges contacting the ground, supporting the weight on the knuckles. Gorillas and chimpanzees are habitual knuckle-walkers, whereas orangutans usually move quadrupedally with the hand made into a fist. IM indexes for the chimpanzee, gorilla, and orangutan are 102, 116, and 139, respectively.
6. Leg longer than arm - leaping and arboreal
A special class of leaping locomotor behavior, in which the body is positioned vertically at rest, is called vertical clinging and leaping. It requires powerful hind limbs to propel the leap as well as to break the impact of landing. Most (but not all) vertical clinging and leaping species have a tail that is used maintain attitude control during leaps. Rapid movements are so well-coordinated during flight that the animal transits the crown of a tree without appearing to make contact with branches. This visual impression of suspending the laws of gravity fueled many "ghost" myths associated with vertical clinging and leaping primates. There is a tendency toward elongation of tarsal elements, especially calcaneus and navicular. Posterior elongation of tuberosity of the calcaneus serves as a robust lever arm for M. gastrocnemius and M. soleus, powerful flexors of the foot. The tendency for fusion of the tibia and fibula is fully expressed only in the tarsier. IM index is below 70.
7. Leg longer than arm - bipedalism
Though obligate bipedalism is found only among humans, many other primate species are capable of facultative bipedalism. Foot specializations for bipedalism include an enlarged and robust tarsal region, greatly reduced phalanges, and strong ligaments that bind tarsals and metatarsals into shock-absorbing longitudinal and transverse plantar arches. A large calcaneus tuberosity acts as a lever arm for plantar flexion. The most unique character of the long, robust legs is the placement of the knees (when in anatomical position) close to the median sagittal plane, functionally beneath the body's center of gravity. The knee itself is adapted to locking in full extension with deep groves to stabilize the patella, a bone that forms in tendons of the quadriceps muscle. The broadened hip becomes a primary weight-bearing joint, characterized by an enlarged femur head as a weight-bearing surface. Pelvic anatomy is dramatically rearranged. A relatively broad sacrum positioned above the hip joint transfers weight to femur head via a wide and robust illium. A shortened ischium places the ischial tuberosity relatively close to the acetabulum. The vertebrae, increasing in size progressively from skull to sacrum, are arranged in a ventral-dorsal S-shaped curve above the pelvis. Though free of locomotor tasks, the arm retains the range of movement seen in brachiators. IM index is 70.
8. Climbing by nails
Elongated and laterally compressed nails of callithricines have the functional attributes of claws. Although they climb by grasping small branches, they are able to use these specialized nails to cling to relatively flat, vertical surfaces of larger trees. IM index ranges from 70 to 80
Color vision is a primate characteristic that presumably reflects our arboreal ancestry. There two types of photoreceptors in the primate retina: rods which function better at low light levels (scotopic vision), and cones which respond to much higher light intensities (photopic vision). The eyes of most diurnal mammals have cones more numerous toward the center of the retina (the region of sharp focus) and more rods toward the periphery. Nocturnal primates have only rod photoreceptors in the retina. The retina of higher primates has a macula lutea (yellow spot) of cones. The fovea, a small depression in the center of the macula in which there is only a single layer of cones, is the area of keenest vision and the target of focusing by the lens.
Color vision is produced by photosensitive pigments that differentially absorb wavelengths. Color perception depends upon the relative degree to which each pigment is stimulated. Primates have three different pigments, producing trichromatic vision.
Anthropoid vision is stereoscopic; that is, the eyes are positioned forward, allowing an overlap of most of the fields of vision with the optic axes parallel. An object is focused on both retinas simultaneously. The optic nerve tracts that pass information from retina to the brain meet at the optic chiasma. In most vertebrates and marsupials the fibres of the optic nerves cross at the chiasma and pass to the opposite of the brain. However in mammals, some of the fibres do not cross over so information from each eye is processed on both hemispheres of the brain
Appendix B: More on Differences
"I'm not interested in what I share with the mouse; I'm interested in how I differ from our closest relatives, chimpanzees," says Svante Pððbo, a geneticist at the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany. Such comparisons, he argues, are the only way to understand "the genetic underpinnings of what makes humans human." With the human genome virtually in hand, many researchers are now beginning to make those comparisons. At a meeting here last month, Pððbo presented work by his team based on samples of three kinds of tissue, brain cortex, liver, and blood from humans, chimps, and rhesus macaques. Pððbo and his colleagues pooled messenger RNA from individuals within each species to get rid of intraspecies variation and ran the samples through a microarray filter carrying 20,000 human cDNAs to determine the level of gene expression. The researchers identified 165 genes that showed significant differences between at least two of the three species, and in at least one type of tissue. The brain contained the greatest percentage of such genes, about 1.3%. It also produced the clearest evidence of what may separate humans from other primates. Gene expression in liver and blood tissue is very similar in chimps and humans, and markedly different from that in rhesus macaques. But the picture is quite different for the cerebral cortex. "In the brain, the expression profiles of the chimps and macaques are actually more similar to each other than to humans," Pððbo said at the workshop. The analysis shows that the human brain has undergone three to four times the amount of change in genes and expression levels than the chimpanzee brain ... "Among these three tissues, it seems that the brain is really special in that humans have accelerated patterns of gene activity," Pððbo says." (from http://www.sciencemag.org/cgi/content/full/292/5514/44)
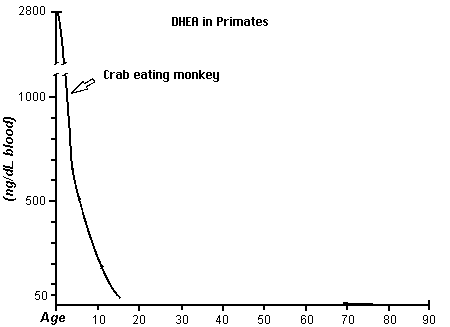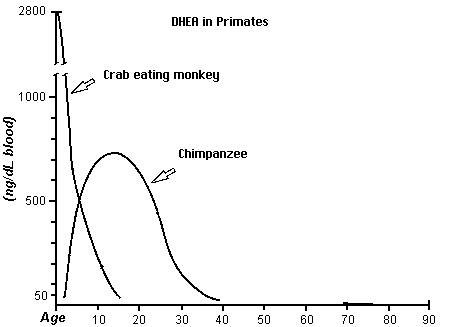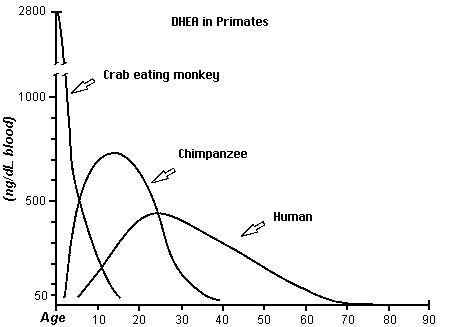
Another known chemical difference is the use of DHEA through out the longer life span of the human as compared to the chimpanzee and a monkey. The use of DHEA may be a main factor in the growth of the brain. In the competition for DHEA among tissues, nervous tissues capture DHEA better than other tissues. Brain tissue naturally contains 6.5 times more DHEA than is found in other tissues. Also DHEA along with testosterone may be a factor in hair growth.
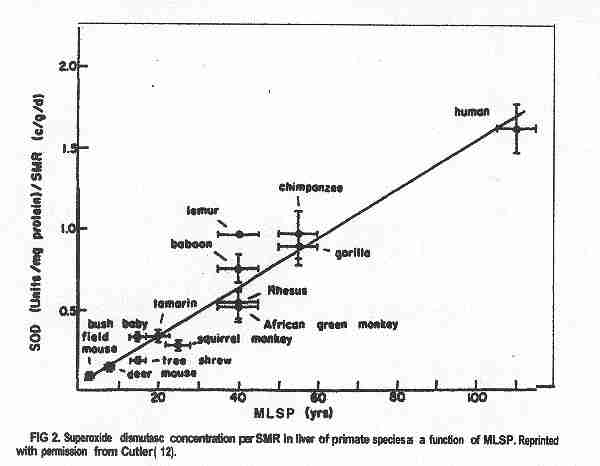
This figure is derived from the first chart (human DHEA); Journal of Reproduction and Fertility 1985; 74: 347, from Text- fig. 5, page 355 (monkey); and J. Repro. Fert. Supplement No. 28; 1980, from Text-fig. 5, page 137 (chimpanzee) (from James Michael Howard http://www.anthropogeny.com)Studies of the usage of the antioxidants have revealed another difference. Antioxidants may be a major factor in health and longevity.
The studies will continue as the evolutionists search for the ultimate proof that man and chimps evolved from a common ancestor!! But, as more and more differences are discovered, which is proved, the theory of evolution, or the creation record of the Scriptures?
Some keys: chimpanzee, chimps, brain, human brain, primate brain, ape brain, chimpanzee brain, brain size, large brain, brain function, language, language development, speech, speech mutation, speech anatomy, speech center, eyes, communicative eyes, beautiful eyes, expressive eyes, white of the eyes, human body, human mutation, human anatomy, foot, foot evolution, bipedal foot, foot structure, foot print, primate foot, chimpanzee foot, ape foot, foot mutation, foot size, foot pattern, brain stem, brain waves, brain rhythm, brain membrane, brain parts, brain function, brain dead, brain cavity, brain image, vision .
